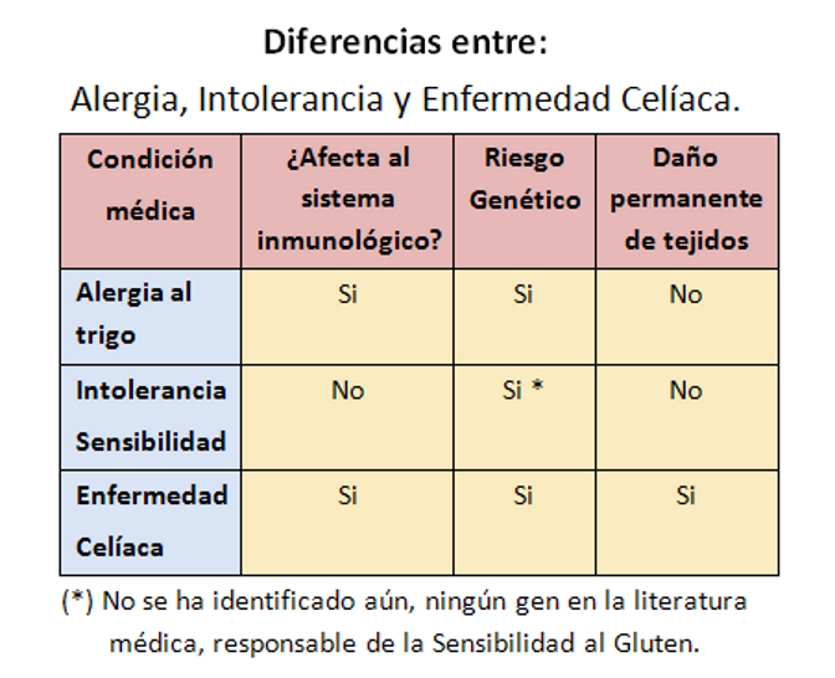
 Los trastornos relacionados con el gluten, se dividen en 3 formas de reacción al gluten:
Los trastornos relacionados con el gluten, se dividen en 3 formas de reacción al gluten:
1) alérgicas: alergia respiratoria, alimentaria, asma ocupacional y urticaria o anafilaxis inducida por el ejercicio;
Las reacciones alérgicas se inician en minutos a horas post exposición al gluten y los anticuerpos IgE juegan un papel central en la patogénesis. Su determinación, junto con los tests de reactividad cutáneos, constituyen pruebas útiles para su diagnóstico.
2) autoinmunes: enfermedad celíaca (EC), dermatitis herpetiforme (DH) y gluten ataxia (GA);
Las reacciones autoinmunes, se desarrollan en tiempos, que van desde semanas, hasta años post exposición del gluten. La más conocida de estas afecciones es la Enfermedad Celíaca, definida como una enteropatía autoinmune, desencadenada por ingesta de gluten en individuos genéticamente susceptibles. Afecta el 1% de la población y su frecuencia está en aumento, quizás debido a la ‘occidentalización’ de la dieta.
Las formas clínicas incluyen, la clásica, la extradigestiva, la silente y la potencial En el diagnóstico se utilizan marcadores serológicos: IgA anti-transglutaminasa tisular (Ac. TG-t) (test inicial); IgA antiendomisio (test confirmatorio); y anticuerpos anti péptidos deamidados de la gliadina (particularmente útiles en deficiencias de IgA y niños menores de 3 años) y las biopsias de intestino. Para su diagnóstico se ha sugerido recientemente, que basta con la presencia de 3 o 4 de los siguientes criterios: clínica típica, serología específica de clase IgA en títulos altos, genética, enteropatía en la biopsia y respuesta a la dieta libre de gluten (DLG).
Otra forma de enfermedad autoinmune vinculada al gluten es la Dermatitis Herpetiforme, consiste en la aparición máculas, luego pápulas y vesículas que dejan costras, pruriginosas, simétricas y de curso crónico.
La erupción de la DH por lo general es simétrica y en la mayoría de los casos compromete codos y partes extensoras de brazos, seguido de compromiso de glúteos y rodillas,, parte superior de la espalda, posterior del cuello y cuero cabelludo. También se afecta frecuentemente el rostro y la línea de implantación del pelo. Las lesiones orales, palmares y plantares se han considerado lesiones raras en esta enfermedad, aunque algunos autores reportan lesiones orales en más del 70% de los casos en pacientes con DH activa. Los pacientes con DH y enfermedad celiaca tienen sintomatología oral más frecuente que los pacientes con DH sin enfermedad celiaca.
El depósito de IgA granular en la dermis papilar en la biopsia de piel sana y la serología positiva para EC (análisis de Anticuerpos Antitransglutaminasa y Antiendomisio), permiten el diagnóstico sin necesidad de realizar la 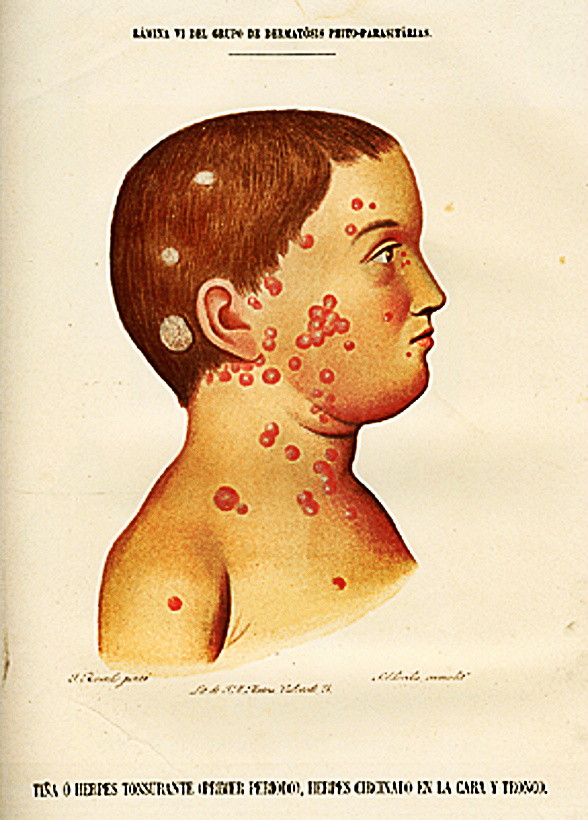 biospia de intestino.
biospia de intestino.
El tratamiento con dapsona es efectivo para la enfermedad y asociado a una dieta libre de gluten se disminuyen los requerimientos del medicamento.
La edad de inicio de esta enfermedad tiene un amplio rango que va desde los 10 meses a los 90 años; sin embargo,es rara en niños, aunque si ocurre se ve más entre los 2 y los 7 años.
La edad promedio de inicio de la Dermatitis Herpetiforme es a los 40 años, a diferencia de la enfermedad celiaca que se inicia más frecuentemente en la niñez, aunque también puede iniciarse a cualquier edad. Se presenta con mayor frecuencia en hombres que en mujeres con una proporción de 1.5:1 aunque la enfermedad celiaca es predominante en mujeres. La prevalencia de las variaciones regionales es muyimportante ya que se deben sospechar factores etiológicos como variaciones en la ingesta de gluten y yodo, aunque algunos estudios no han mostrado que la ingesta de yodo afecte la prevalencia. La incidencia de la DH es más frecuente en británicos y europeos del norte posiblemente por el bagaje étnico; los factores estacionales y de temperatura no se han correlacionado con la actividad de la enfermedad.
La tercera forma de reacción autoinmune es la “ataxia asociada al gluten” o gluten ataxia, la cual consiste en una forma esporádica de ataxia cerebelosa pura, de comienzo insidioso en la edad media de la vida que tiene marcadores serológicos de sensibilización gluten.
y 3) sensibilidad al gluten no celíaca (SGNC): en la que no hay evidencia de reacciones autoinmunes ni alérgicas pero probablemente sea inmuno-mediada.
Ésta forma inmuno-mediada de intolerancia al gluten se inicia horas a días luego de la exposición y se la denomina “sensibilidad al gluten”. En esta condición los pacientes refieren sentirse mal cuando ingieren gluten y mejorar al excluirlo de la dieta, habiendo sido convenientemente excluidos los diagnósticos de alergia al gluten y EC. También se debe haber descartado la deficiencia de IgA.
![]()